Wiedza na temat struktury trójwymiarowej lipaz oraz czynników, które określają ich regiospecyficzność i enancjospecyficzność są istotne przy wyszukiwaniu zastosowań przemysłowych dla tych enzymów. Lipazy HPL oraz RML były pierwszymi opisanymi były pierwszymi opisanymi, dzięki analizie z wykorzystaniem promieni X, strukturami lipaz. Następnie poznano wiele różnych struktur lipaz pochodzenia grzybowego od: GCL, FSL, CRL [63], CAL, HLL i Rhizopus delemar (RDL) [20;34]. Struktury lipaz bakteryjnych poznawane były dużo wolniej. Pierwszą była lipaza od Bacillus glumae (BGL) w 1993 roku, następnie kilka lat później lipaza CVL, (lipaza, która wydawała się być identyczna jak lipazy BGL i BCL). Stosunkowo niedawno opublikowano struktury lipaz pochodzących od SEL, Streptomyces scabies (SSL) i esterazy Pseudomonas fluorescens [66]. Ten ostatni enzym może hydrolizować triacetynę i tributyrin, ale nie może hydrolizować triglicerydów długołańcuchowych kwasów tłuszczowych [33;67].
Na podstawie pierwszych trójwymiarowych struktur lipaz HPL i RML odkryto, że zawierają one katalityczną triadę Ser-Asp-His podobną do triady proteaz serynowych [77]. Triada katalityczna jest miejscem aktywnym α/β fałdowych hydrolaz i zawiera trzy katalityczne reszty, występujące zawsze w tej kolejności w sekwencji aminokwasowej (rys.3):
Kolejność ta jest odmienna od zaobserwowanej wcześniej u innych białek, które zawierają triadę katalityczną. W lipazach nukleofilowych zawsze uważano, że występuje reszta serynowa, podczas gdy katalitycznym kwasem jest reszta asparaginowa lub glutaminowa [33].
Nukleofilowa reszta serynowa została zlokalizowana w wysoce zakonserwowanym pentapeptydzie Gly - X - Ser - X - Gly,(X - dowolny aminokwas) który tworzy ostry, γ - podobny zakręt pomiędzy β 5 centralnej β - kartki kanonicznej α/β hydrolazy i następującej α helisy. Zakręt γ jest określony przez niekorzystny energetycznie główny łańcuch φ i kąt skręcenia reszty nukleofilowej. β 5, γ-zakręt i następująca α - helisa tworzą najlepiej zakonserwowaną strukturalną cechę α/β fałdowych hydrolaz. Ponieważ istnieją bliskie kontakty pomiędzy resztami, dwie pozycje przed i dwie pozycje za resztą nukleofilową, w jednej lub zwykle dwóch pozycjach występuje glicyna (Gly). Jest ona czasami podstawiana przez inne mniejsze reszty, takie jak alanina (Ala), walina (Val) lub treonina (Thr). Przykładem może być lipaza Bacillus subtilis (BSL) posiadająca sekwencję Ala - X - Ser - X - Gly dookoła miejsca aktywnego seryny [67]. Rozmieszczenie skrętu nukleofilowej helisy zostało nazwane "nukleofilowym zakrętem" (ang. nucleophile elbow). Zgodna sekwencja lipaz Ala / Gly - X - Ser - X - Gly faktycznie może zostać wyjaśniona przez przestrzenne wymagania nukleofilowego zakrętu. Cała katalityczna triada znajduje się w hydrofobowym otoczeniu. Seryna jest pasmowej konformacji, z niekorzystnymi kątami skrętu (β - ε Ser - α) i znajduje się na hydrofobowej powierzchni. Jednakże, jako wynik tej unikalnej architektury, grupa hydroksylowa seryny posiada dostęp do katalitycznej histydyny i atomu węgla grupy karbonylowej wiążącego cząsteczkę substratu [67]. Zakręt nukleofilowy pozycjonuje zatem wolne reszty nukleofilowe powierzchni miejsca aktywnego i pozwala na łatwy dostęp do miejsca aktywnego, z jednej strony dla reszty histydyny, a z drugiej zaś dla substratu. Ostry zakręt również optymalnie pozycjonuje resztę nukleofilową N - końca kolejnej helisy, tym samym pozwalając na stabilizację tetraedrycznego produktu pośredniego i zjonizowanej formy reszty nukleofilowej [33]. Jak wspomniano wcześniej, hydrolazy acyloestrowe są funkcjonalnymi analogami proteaz serynowych. Umieszczenie reszt katalitycznych triady wykazało, że aktywny atom nukleofilu (Ser-Oγ) i odpowiadająca grupa kwasu karboksylowego (Asp lub Glu) znajdują się w jednej płaszczyźnie z histydyną imidiazolową w obydwu rodzinach enzymu, tworząc sieć podobnych wiązań wodorowych, która jest niezbędna dla procesu katalizy. Poza zgodnym serynowym miejscem aktywnym, istnieje bardzo niewielka zgodność sekwencji pomiędzy różnymi lipazami, z wyjątkiem enzymów pochodzących od blisko spokrewnionych źródeł [67].
U prototypowych α/β fałdowych hydrolaz, kwas katalityczny (Asp lub Glu) występuje po odwrotnej stronie zwoju po paśmie 7 centralnej β - kartki. Jest nim aktywne miejsce histydyny połączone wiązaniem wodorowym. Jednakże, typologiczna pozycja kwasu katalitycznego wydaje się być zmienna: w przypadku lipazy trzustkowej występuje za pasmem 6. Obserwacja ta zachęciła Schrag [64] do zmiany konstrukcji w lipazie GCL, przez zmianę pozycji kwasu katalitycznego z pasma 7 na 6. Podwójny mutant zachował ok. 10% pierwotnej aktywności, potwierdzając różnorodność pozycji kwasu katalitycznego. Podobnie, funkcjonalność kwasu katalitycznego w przypadku dehalogenazy (Asp - 260 występującej po pasmie 7) może być uwolniona dzięki mutacji Asn - 148Asp, reszty ulokowanej za pasmem 6. Struktura otrzymana dzięki analizie z użyciem promieni X lipazy BGL ujawniła Asp - 263 jako kwasową resztę katalityczną, która jest połączona wiązaniem wodorowym z katalityczną histydyną. Jej pozycja nie występuje na końcu β 7. Pomiędzy β 7 i Asp - 263 leży kilka elementów o strukturze drugorzędowej. Mutagenezy centrum aktywnego wykazały, jednakże, redukcję aktywności mutanta Asp - 263Ala, ponieważ grupa karboksylowa Glu - 287, która występuje po paśmie 8, leży blisko katalitycznej histydyny, zrozumiałym jest, że Glu - 287 przejmuje rolę Asp - 263, każda bezpośrednio lub poprzez cząsteczkę wody.
Trzecim wiązaniem katalitycznym w lipazach jest wiązanie poprzez katalityczną histydynę. Reszta ta jest ulokowana w pętli po paśmie β 8 α/β fałdowej hydrolazy. Długość i konformacja tej pętli są różne.
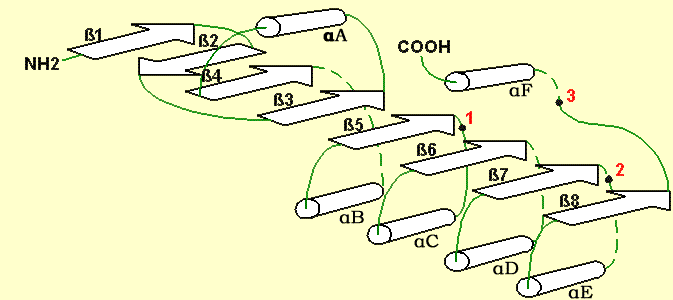
Rys.3 Ulokowanie katalitycznych reszt miejsca aktywnego α/β fałdowych hydrolaz: (1) - nukleofilowa reszta serynowa (Ser, Cys, Asp); (2) - reszta kwasowa (Asp lub Glu); reszta histydynowa (His) [33].
Nałożenie struktur lipaz, w połączeniu z wyrównaniem sekwencji, zakonserwowanych reszt, które powodują utrzymywanie struktury białka, ujawniło, że poza miejscem aktywnym, większość niezmiennych reszt jest ulokowanych na brzegach drugorzędowych elementów strukturalnych, tj. mostków disiarczkowych lub solnych, oraz w rdzeniu białek. Z drugiej strony wysoką różnorodność pozycji znaleziono na powierzchni enzymów i w sąsiedztwie miejsc wiązania substratu. Zakłada się, że katalityczna triada (Ser - Asp / Glu - His), podtrzymywana na centralnej ß-kartce z podobną drugorzędową strukturą wokół aktywnego miejsca seryny (ß - ε Ser - α), jest wspólnym motywem we wszystkich seryno - zależnych lipaz i esteraz. Od czasu, gdy porównywalna topologia centralnej katalitycznej domeny została zauważona w pozornie różnych niespokrewnionych hydrolazach, zasugerowano, że wszystkie enzymy o α/β fałdowaniu hydrolaz pochodzą od wspólnego przodka. Ewolucja rozbieżna, która zakonserwowała strukturalny szkielet kluczowych katalitycznych komponentów, przy braku całkowitych podobieństw sekwencji pomiędzy wyraźnie spokrewnionymi enzymami jak lipazy i esterazy, została połączona z domenami określającymi różne specyficzności [67].